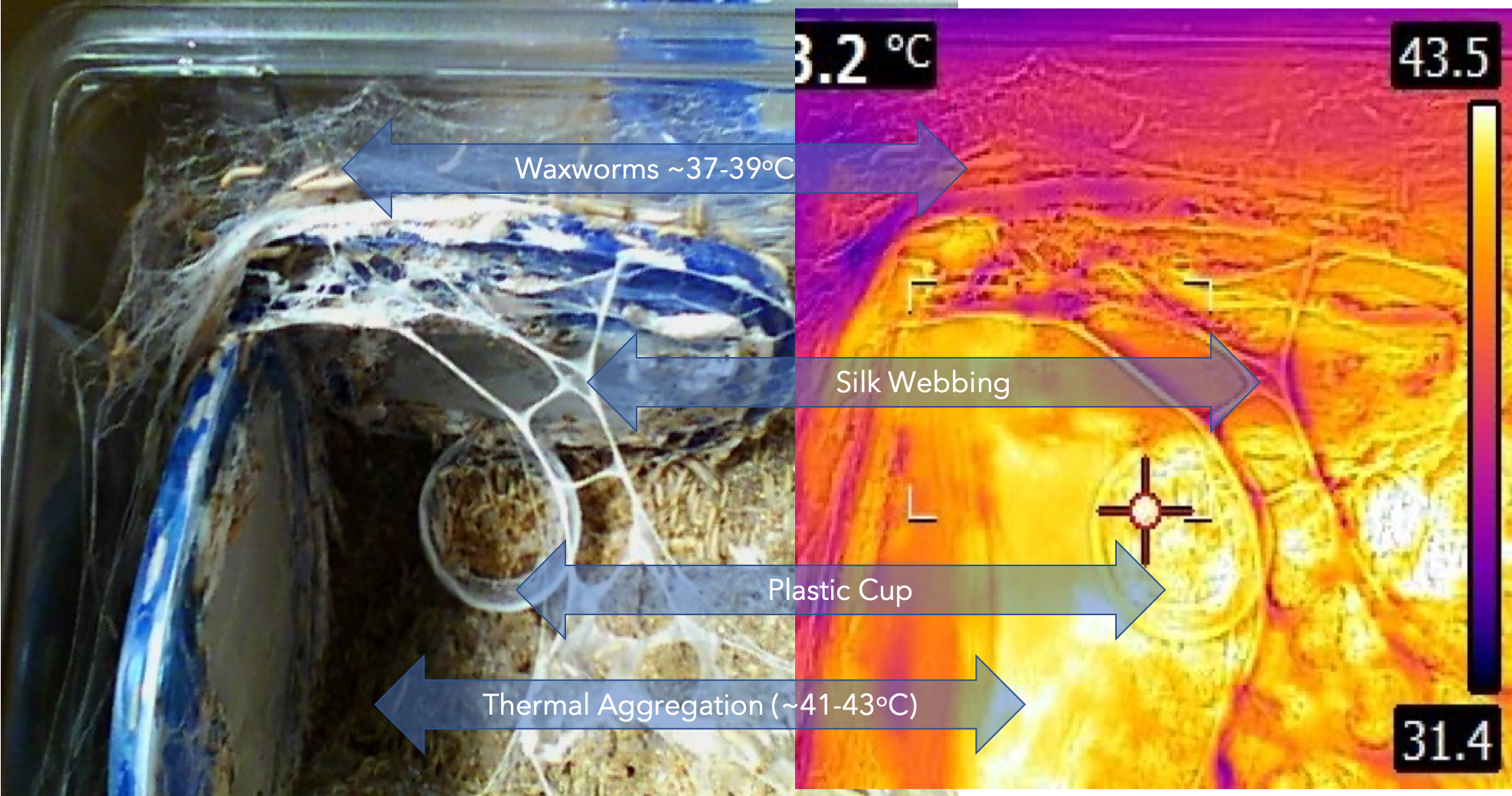
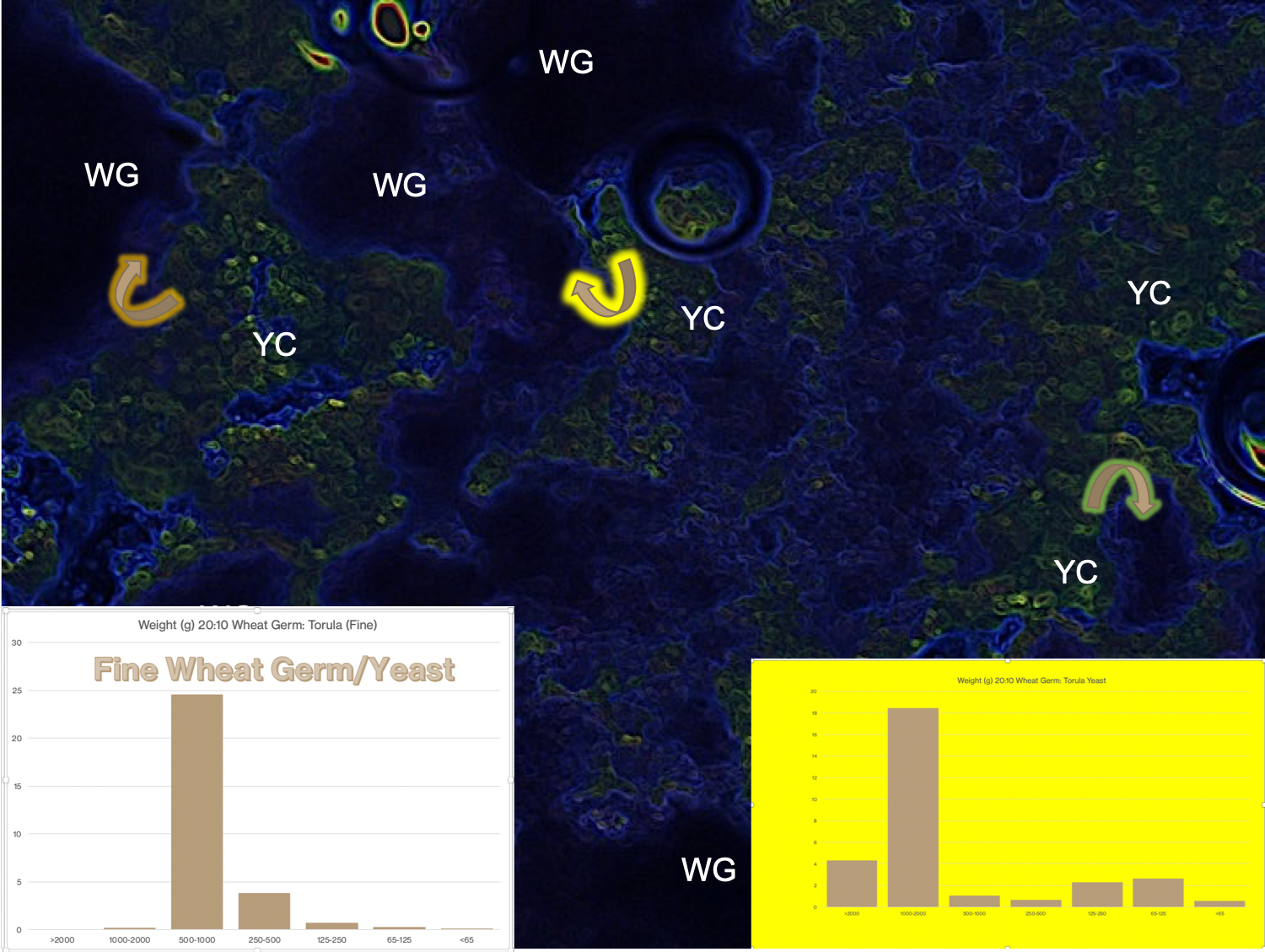
In a recent class we were discussing soy flour & other soy products in our insects’ diets. A student expressed confusion about the term “ANTI-NUTRIENT.” The student suspected that something in her insects’ diet might be causing the unreliable outcomes in her rearing. After reviewing the diet components, the soy flour raised suspicion, as the student’s rearing facility had to use several brands of soy flour. This raised the question and ensuing dialog about the benefits and liabilities of soy in diets, including the ANTI-NUTRIENT concept along with the factors that are assets of soy (high protein content, beneficial lipids, antioxidants, etc.) The out-of-class discussion (by email) led to Cohen’s provision of the image above, summarising the anti-nutrient factors, and after in class discussion of this issue, Professor Cohen realised that the soy benefits and liabilities questions would benefit the whole class as to the concept of “ambiguous” potential of soy in diets. This led to a whole hour of discussion on biological activities of soy, and to support and enhance this new topic, Cohen provided the link for the below document by Michio Kurosu: Biologically Active Molecules from Soybeans.
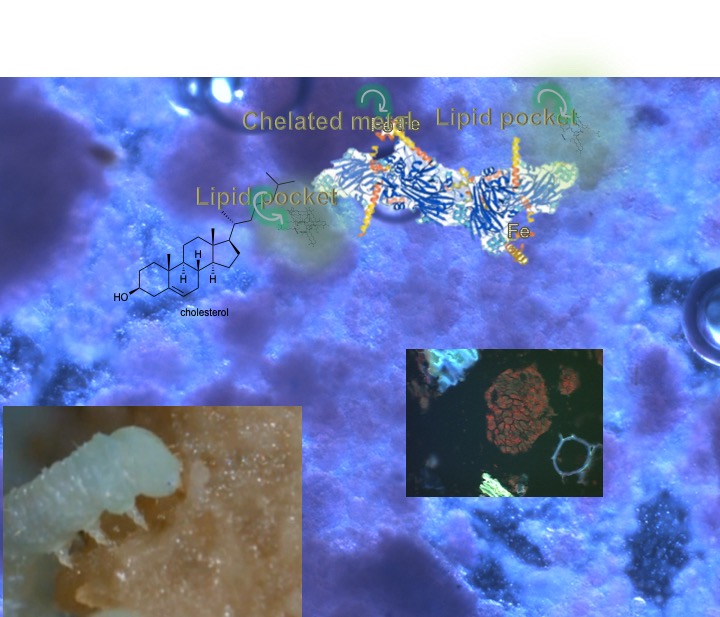
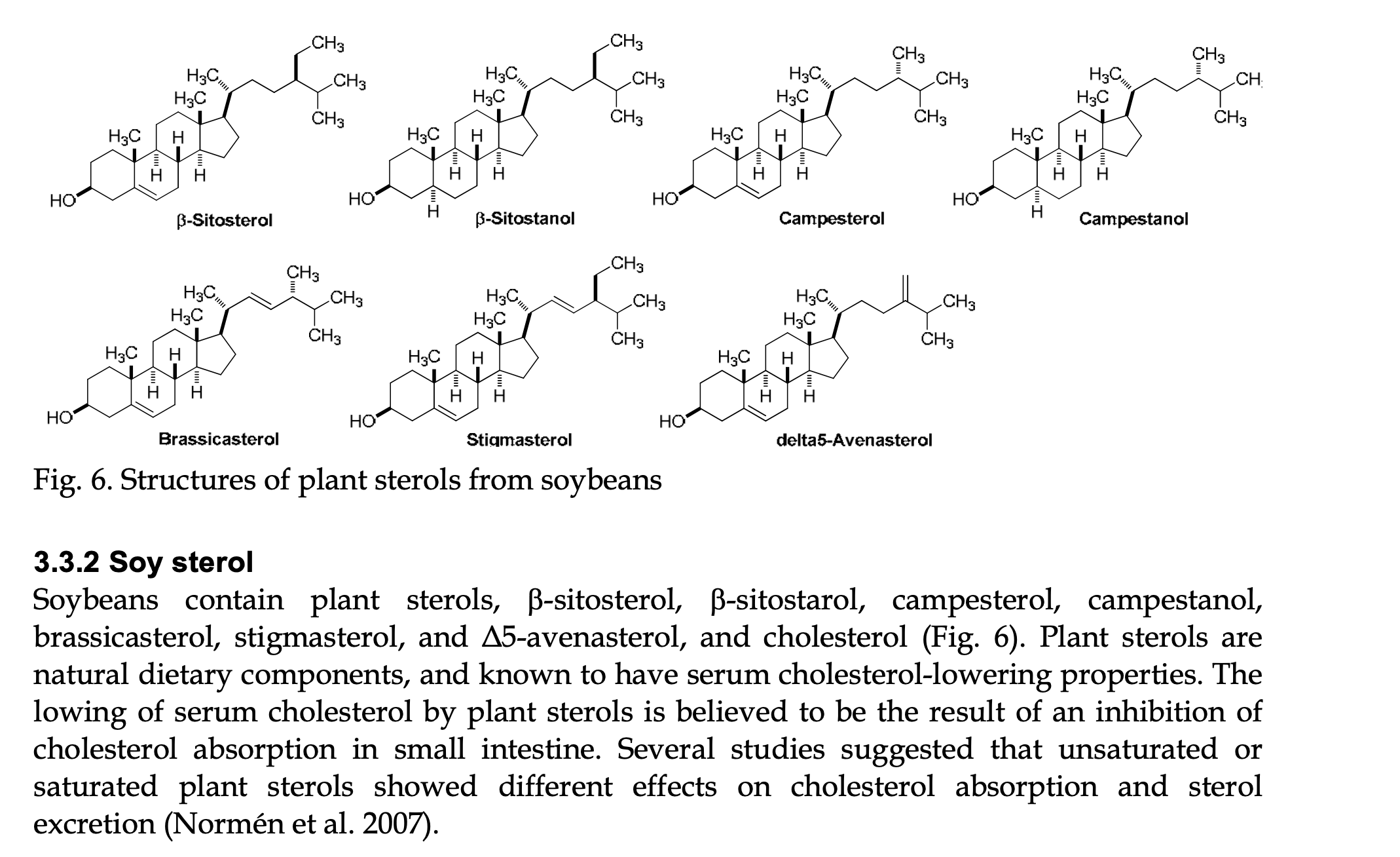
This amazing paper (available through OPEN ACCESS) contains a wealth of information about soy molecules, many of which are of tremendous relevance to insect diets. As an example, I have provided a screenshot of a page that shows the structures and names of the phytosterols found in soy. This content led to further discussion in class (with several conferences by email after class) on how soy has “hidden” benefits that may not have been primary reasons for selecting soy but which are worth considering in our every-present desire to engineer our insects’ diets to be more successful.
The bottom line of all this is the value of the way Allen Carson Cohen teaches his courses. This kind of enrichment is and ongoing process in Cohen’s continual quest to better understand the mechanisms of insect diets and other aspects of insect rearing systems.
I (Allen Carson Cohen) intend to spend the next several weeks providing information about this comprehensive, “deep inquiry” approach to insect rearing. I hope that readers will provide feedback about this–hopefully informative–journeys into rearing science.