Allen Carson Cohen teaching a recent Fundamentals of Insect Rearing class (Figure 1 for Feb. 21, 2026 post).
SOME BIOGRAPHICAL ANECDOTES BEHIND MY APPROACH TO INSECT REARING EDUCATION. Hi insect rearing community! Though I have interacted with many of you from the rearing community as colleagues, students, and others, I decided to provide some background that explains my current involvement in insect rearing.
I started my formal professional work with insect rearing as an older post-doc at the University of Arizona and the USDA, Agricultural Research Service in 1979. Prior to accepting that position, I had been teaching biology at Cypress Community College, in Cypress, California for over a decade, and I did my doctorate in entomology at UC, Riverside during my Cypress teaching phase. The job at U of A and at the USDA was to develop an artificial diet for predators to be used in a mass-rearing program to control crop pests, mainly in cotton and alfalfa cropping systems. At the start of my post-doc, I knew nothing of insect rearing, as my background was with field-collected blister beetles (Family: Meloidae). Because I had some background in physiological and biochemical ecology of insects, Drs. George Ware and Harry Graham thought I might find my way through the complexities of diet development.

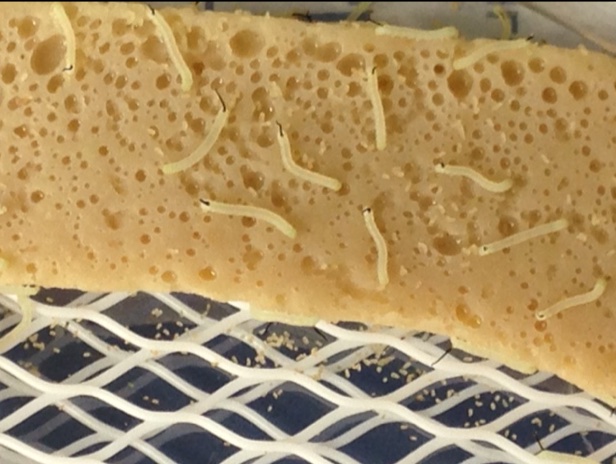
Figure 2 (Feb. 21, 2026 post). Big-eyed (Geocoris punctipes) bug eating a cotton aphid (Aphis gossipii) (picture taken by Jack Dykinga in Cohen’s USDA, Western Cotton Lab, Phoenix, an ARS lab, circa 1995). Please note that this image is part of the rearing course I deliver online (Zoom) several times a year.
The picture of the big-eyed bug eating an aphid typifies what I was seeing in the first years of my efforts to develop an artificial diet that would support a program of mass rearing of this or other predators. Seeing the piercing-sucking mouthparts of the big-eyed bug penetrating the aphid, I assumed (and was informed by the then current literature) that the hemipteran predator was inserting a kind of drinking straw into the prey and sucking out the fluids (hemolymph) of that prey.
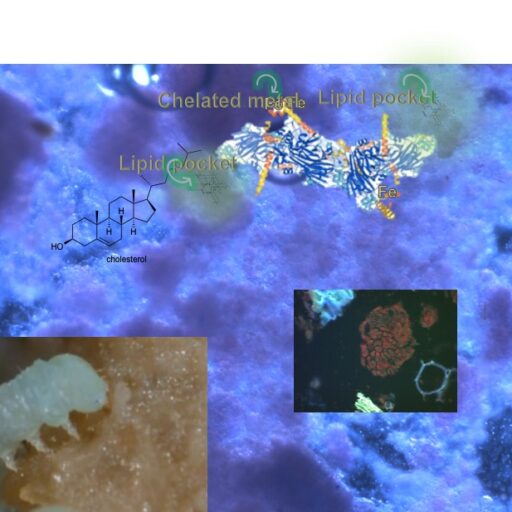
For nearly 3.5 years, I struggled with efforts to design a liquid diet that would match the hemolymph chemistry of various prey of the hemipteran predators (beet army worms, corn earworms, aphids, etc.) I used gas chromatography, HPLC, amino acid analyzers (ion exchange systems), atomic absorption spectroscopy, etc. to analyze the contents of hemolymph and then I would try to formulate liquids that reflected the lipids, amino acids, carbohydrates, etc. of prey hemolymph, and I struggled further to develop a liquid diet presentation system that would simulate the cuticle and body configuration of typical prey. ALL THESE EFFORTS WERE OF NO AVAIL!
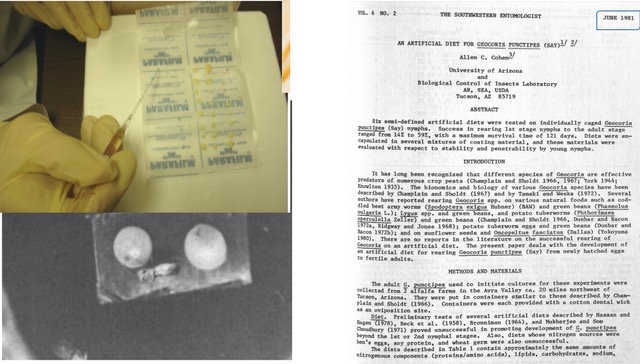
Figure 3 (Feb. 21, 2026 post) illustrates the system I used to provide a coated liquid diet for my big-eyed bugs (left side). It also shows an interim paper that I wrote as a progress (or lack of progress) report on liquid diets for G. puncitpes.
SUCCESS AT LAST! It was not until I started studying the true feeding habits of big-eyed bugs and their hemipteran (heteropteran) cohorts that I was able to apply the basic knowledge I gained about extra-oral digestion to develop a solid/semisolid artificial diet for G. puncitpes and, in turn, for other arthropods.
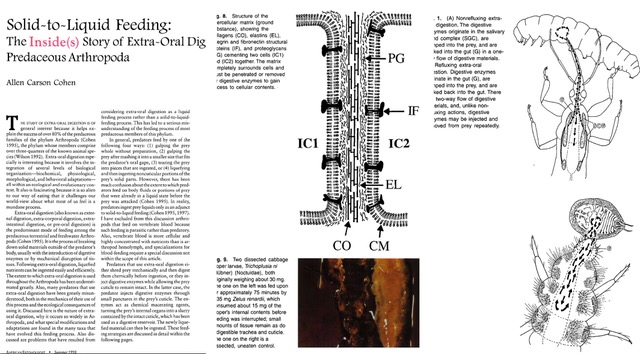
Figure 4 (Feb. 21, 2026 post) showing the strides that I had made in diet development (practical accomplishments) and the deeper understanding of extra-oral digestion (= EOD) in basic insect science (biochemistry, morphology, behavior, ecology, physiology). The 1998 paper (on the left) represents the publication on EOD, which I called “Solid-to-Liquid Feeding: The Inside(s) Story of Extra-Oral Digestion in Predaceous Arthropods. The images on the right side of Figure 4 show the kinds of extra-oral digestion (what I called non-refluxing and refluxing feeding), and the middle images show the structure of extracellular matrix that holds together the cells of the prey. The paper was a feature article in a 1998 American Entomologist.
will leave off here for today’s blog post, but I will take up from here with some specifics about how and why I developed my approaches to rearing education, including the reasoning behind rationale-driven, basic science-based inquiry. You will soon see how and why my emphasis on learning cause-and-effect relationships, rearing systems interactions, and mechanism-based inquiry offers a high likelihood of success in developing new rearing systems, improving (optimizing) existing systems, and developing data-driven standard operating procedures (SOPs) for rearing systems.