To view the video in this tutorial example, please right click the image and select “Show Controls,” then click the forward arrow to view the video.
The special feature of this approach is that collaborators or teacher/learners can communicate live through Zoom or other remote formats and share ideas, explanations, etc.
This is an example of a tutorial where the collaborators (Professor Cohen and Dr. Carole Cheah) were exchanging information about diet development for their project dedicated to development of artificial diets for predators that are specialists of hemlock woolly adelgids. In this video, recorded from a video collaboration, Drs. Cheah and Cohen review various aspects of the acceptance of the artificial prey (artificial diet later published in a 2015 paper (published here: Cohen and Cheah, Entomol Ornithol Herpetol 2015, 4:2 http://dx.doi.org/10.4172/2161-098)
The main point of this blog page is to illustrate the potential quality and efficiency of remote communication (Cheah was in Connecticut and Cohen was in North Carolina for this Zoom meeting). Note that the video here includes only Dr. Cheah’s part of the communication where Cheah showed Cohen videos and still shots of the beetles she was feeding diets produced by Cohen as freeze dried samples of their “CC Diet.” Dr. Cheah was testing the responses of the beetles to the diets with various additives and diet presentation conditions such as on or off of hemlock foliage or Parafilm or other substrates. Please note further that Dr. Cheah at the time was using a simple phone-camera and shot the videos and stills through a dissecting scope eyepiece.
This type of exchange works well for many other kinds of information exchanges which can be done live and which can further be made to include multiple research or rearing education partners.
Cohen is currently offering this type of collaboration and teaching as part of an online experience for collaborators or participants worldwide.
This post and several others to follow are intended to narrate how I approach teaching about insect rearing systems and how I approach developing and improving these systems. I am writing this page on February 14, 2025 almost 50 years after I started out in insect rearing at the University of Arizona and the USDA, Agricultural Research Service in August of 1979. During this time, I have had several successes and several failures at developing and/or improving rearing systems. It has become the focus of my life to better understand why systems and the components of those systems work or fail. These pages are my effort to explain what I have learned in trying to understand the scientific principles behind rearing.
First, the concept of the rearing system as an ecological niche: before I was an entomologist/rearing specialist, I was an ecologist. Over the years, I have come to appreciate the reality that as rearing specialists, we are obligated to provide all the components that our insects would have in nature–only we must put them all into our rearing system. THIS IS TO SAY THAT OUR REARING SYSTEMS ARE ARTIFICIAL ECOLOGICAL NICHES!
As “NICHE-KEEPERS,” we must provide the food, the gas exchange, the heat, light, humidity, sites for oviposition, microbial relations, mating sites, oviposition sites, etc., etc. Are we giving our insects the right conditions to safely void their urine and fecal wastes? You watch a leaf-eating caterpillar in nature dropping its frass from the leaf-feeding site to the ground. How must they release their wastes in our containers? We know what is convenient for us as the rearing personnel, but is that what is best for the insects?
The two pictures in this post show a little of what I mean by factors in the ecological niche concept of insect rearing systems. The top picture is an attempt for me to show in multiple dimensions (based on G. Evelyn Hutchinson’s “N-Dimensional Hypervolume” as the model of an ecological niche). The idea here is that CO2 generation and O2 uptake are not only related to one-another, but they relate to the thermal conditions and the food the insect is eating. The ratio of CO2/O2 or the respiratory quotient are indicators of whether the insect is metabolising mainly carbohydrates, lipids, or proteins. If being a little confusing with all this and other factors seen in the top diagram, it’s what I intend to do to convince you that this is all pretty complex and VERY INTER-RELATED stuff.
The second picture showing the diet (developed by R.T. Yamamoto 1969) for Manduca sexta is also a complex of components such as wheat germ, casein protein, torula yeast, vitamins, and minerals that are all interacting with one-another to give the diet a composition, texture, consistency, and configuration that help determine the success of our tobacco hornworm rearing system. One reason that the diet is (in my opinion) so successful for M. sexta is that it provides lipids (fatty acids and sterols, for example) in the form of lipoproteins which make these essential lipids bioavailable for the larvae. One of the wonderful features of wheat germ is that it contains nutritious proteins with all the essential amino acids, but also some of these proteins are lipoproteins, which carry the lipids in a bioavailable way compatible with our hornworms’ digestive system.
More along this line in my next blog entry. Happy Valentine’s Day!
In a recent class we were discussing soy flour & other soy products in our insects’ diets. A student expressed confusion about the term “ANTI-NUTRIENT.” The student suspected that something in her insects’ diet might be causing the unreliable outcomes in her rearing. After reviewing the diet components, the soy flour raised suspicion, as the student’s rearing facility had to use several brands of soy flour. This raised the question and ensuing dialog about the benefits and liabilities of soy in diets, including the ANTI-NUTRIENT concept along with the factors that are assets of soy (high protein content, beneficial lipids, antioxidants, etc.) The out-of-class discussion (by email) led to Cohen’s provision of the image above, summarising the anti-nutrient factors, and after in class discussion of this issue, Professor Cohen realised that the soy benefits and liabilities questions would benefit the whole class as to the concept of “ambiguous” potential of soy in diets. This led to a whole hour of discussion on biological activities of soy, and to support and enhance this new topic, Cohen provided the link for the below document by Michio Kurosu: Biologically Active Molecules from Soybeans.
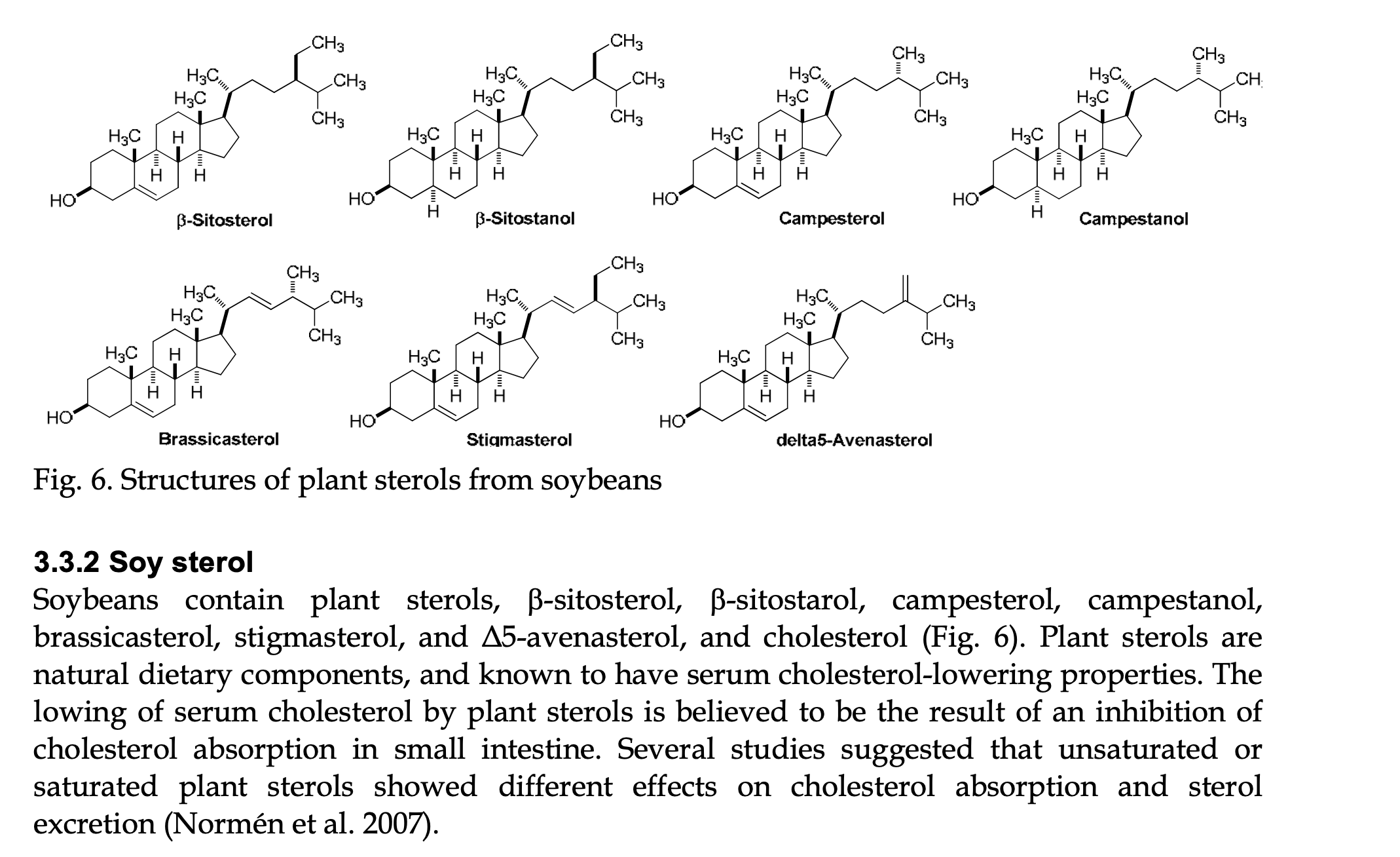
This amazing paper (available through OPEN ACCESS) contains a wealth of information about soy molecules, many of which are of tremendous relevance to insect diets. As an example, I have provided a screenshot of a page that shows the structures and names of the phytosterols found in soy. This content led to further discussion in class (with several conferences by email after class) on how soy has “hidden” benefits that may not have been primary reasons for selecting soy but which are worth considering in our every-present desire to engineer our insects’ diets to be more successful.
The bottom line of all this is the value of the way Allen Carson Cohen teaches his courses. This kind of enrichment is and ongoing process in Cohen’s continual quest to better understand the mechanisms of insect diets and other aspects of insect rearing systems.
I (Allen Carson Cohen) intend to spend the next several weeks providing information about this comprehensive, “deep inquiry” approach to insect rearing. I hope that readers will provide feedback about this–hopefully informative–journeys into rearing science.
Here is an example of how Professor Allen Carson Cohen uses connection with participants to make the educational experience more personalised. Participants in classes view a video or other teaching format and interpret what they are seeing or hearing. They submit (through email or individual Zoom meeting) to Prof. Cohen their interpretation or understanding of what they are seeing, and the instructor replies with comments about how well they have understood the concept being explored.
In this case, students had been told that pH is an important part of how their insects treated natural foods or artificial diets in the insects’ guts. Here, they are seeing the different colours of foods that had been treated with a pH dye that is blue for acids and red for basic pHs. This serves as a basis for an individual/personalized discussion between the participants and the instructor leading to other questions or learning situations that are of special interest to students.
Please note that this format, especially if done in a Zoom meeting or an email with attached images, can allow Professor Cohen to make a virtual visit to your laboratory or workplace to help you more directly with your specific interests. This is what Dr. Cohen calls a “mini-consultation.”
More about virtual meetings and “mini-consultations” in posts to come soon!
We learn best by doing rather than just seeing and hearing. The more we are involved in a learning process, the more we will gain from their educational experience. This axiom is clear to me after decades of teaching, and I keep trying to upgrade my teaching to rise to ever-increasing student/learner involvement.
In Fall 2024, I taught a class in insect rearing for first year life science students here at North Carolina State University.* Here is a sample of one of the slides in a presentation I did on metabolism of waxworms (Galleria mellonella) and how they use lipids and carbohydrates to generate elevated body temperatures.
From this image, FIRST YEAR students were able to write paragraphs explaining their understanding of how metabolism, size (accomplished here by larval aggregations), and insulation (accomplished by the waxworms with silk) could permit what we normally consider “cold-blooded” creatures (insects) to become warm-blooded. What I learned from the interactive paragraphs between the students and myself was that we could conduct a productive discussion that guided the “entry level” students to understand how the concepts behind metabolism, behavioural actions, structural adaptations such as fur, feathers, and silk could lead to waxworms’ cultural characteristics (their success in speeding their development, avoidance of predators and parasites, and many other advantages of making themselves warm.
The main point of this discussion is that the kinds of success we had with extensive student/teacher interactions was a much more efficient and enjoyable learning experience. Having learned this lesson from my freshman students, I vowed to apply it to all my teaching–including my online classes. Please look for future discussions of my interactive teaching ideas in post to come shortly.
We used this image complex in the First Year Life Sciences Class in Insect Rearing (called The Nature of Unnatural Insects: The Science of Insect Rearing Systems). We discussed how the tiny shrew and the world’s smallest hummingbird (each weighing about 2 grams) shared similarities with the waxworms whose thermal signature (lower right, yellow mass) were all able to attain and hold temperatures at least 10 degrees C above ambient. We discussed in our interactive email exchanges how metabolism, structural features such as fur, feathers, or silk could act as insulation, and biomass could be effectively increased by the aggregation of the larvae forming 2 gram masses.
In a recent post (November 30, 2016), I discussed some of the foundations of Drosophila rearing, and I pointed out that much of the rearing that was done over the past century depended on the early works with Drosophila. In fact, the modern concepts of insect rearing must have originated in the 1910 paper by Delcourt and Guyenot: “The Possibility of Studying Certain Diptera in a Defined Environment.” This title, to me, represents and suggests the concept of CONTROLLED REARING.
Baumberger, J. P. 1917a. The food of Drosophila melanogaster Meigen. Proceedings of the National Academy of Sciences of the United States of America: 3: 122-126.
Baumberger, J.P. 1917b. Solid media for rearing Drosophila. American Naturalist. 51: 447-448.
Delcourt, A. and E. Guyenot. 1910. The possibility of studying certain Diptera in a defined environment. Comptes rendus hebdomadaires des séances de l’Académie des sciences (0001-4036), 151, p. 255-257.
Guyenot, E. 1913a. A biological study of a Drosophila ampelophila Low fly I – The possibility of an aseptic life for an individual and the line. Comptes rendus des séances de la Société de biologie et de ses filiales (0037-9026), 74, p. 97-99.
This is a “mini-review” of a new paper that was published on “Genetic and microbiome changes during laboratory adaptation in the key pest Drosophila suzukii. The authors of this paper (K. Nikolouli, H. Colinet, C. Stauffer, and K. Bourtzis) have made this important contribution in the journal Entomologia Generalis, Volume 42 (2022), Issue 5: pp 723-732, and they track wild D. suzukii from the field through multiple generations of laboratory culture. Nikolouli et al. point out that the assumption that field-derived insects make profound changes in their various adaptations, including genetic diversity and symbiotic community changes as they adapt to the artificial conditions of rearing. The authors summarize their work reported in this paper with the following statements “These results can serve as a reference for the design of an area-wide integrated pest management approach with a Sterile Insect Technique (SIT) component. Rearing productivity, biological quality, and mating competitiveness of a SIT mass-reared strain should be assessed in connection with genetic and symbiotic changes occurring during laboratory adaptation.“
The authors statements about the SIT context of these findings actually goes far beyond sterile insect technique, and the findings about changes in genetic characters of the target insects AND the microbiomes have profound implications for rearing for all other purposes, including biological control, insects as food and feed, and research.
A central tenet of the rearing program at NCSU is that insect rearing systems are artificial ecological niches of the insects we rear. This means that every aspect of the insect’s needs must be met by our rearing system. This responsibility of rearing personnel applies whether we know each requirement or not. For example, each insect (and each insect population in our colonies) requires a certain range of oxygen to meet their metabolic needs. We may not know how much oxygen this requires (the range of oxygen concentrations), but when we provide holes in the lid of the rearing container or a screen that permits gas exchange, we hope that the openings are adequate to meet the oxygen demands.
Therefore, we can think of the oxygen requirements as part of our insects’ ecological niche, and if our insects perform adequately under the conditions of our rearing containers, we assume that everything is OK “oxygen-wise.” We make the same kinds of assumptions about carbon dioxide concentrations and air flow and the same about water vapour. Sometimes we get ourselves in trouble when we try to restrict water loss from he diet or the insects by making the openings so limited that we wind up starving our insects of adequate oxygen (a phenomenon known as hypoxia or even anoxia); and/or we make create a situation of excess carbon dioxide, which can threaten our insects’ well-being.
In all the rearing courses that I teach, I try to convey to students that the rearing system with all its components that meet the insect’s ecological niche parameters is a complex set of interactions between various (ALL) components, and a helpful way of viewing all this is with a 3-D model or diagram such as what is seen here:
Model of the complex components in an insect’s ecological and REARING niche.
In this diagram that simulates or suggests three-dimensional space, I have tried to show about 20 of the many, many parameters in a niche. The diagram is intended to illustrate that there are interconnections and interactions that involve all the biotic (biological) and abiotic (physical/chemical) factors that play roles in our insects’ well-being. This model of N-dimensional hyperspace is derived from the work of the famous ecologist G. Evelyn Hutchison. I have used this model in my recent book, Design, Operation, and Control of Insect Rearing Systems 2021, CRC Press, Boca Raton, FL).
For this discussion, let us take-up a simplification of this diagram with 3 factors, heat, CO2, and O2. Here is the simplified diagram:
Three factors (heat, carbon dioxide, and oxygen) that are important in a waxworm rearing system.
In this 3-D, three-factor diagram, I am trying to show that the waxworm from my colony (of Galleria mellonella) is greatly influenced by the heat (temperature) conditions in their rearing container; but also, the concentration of CO2 and O2 are interacting factors that influence the metabolism and aggregation behavior of the waxworms, as well as food consumption, digestion rates, development rates, etc.
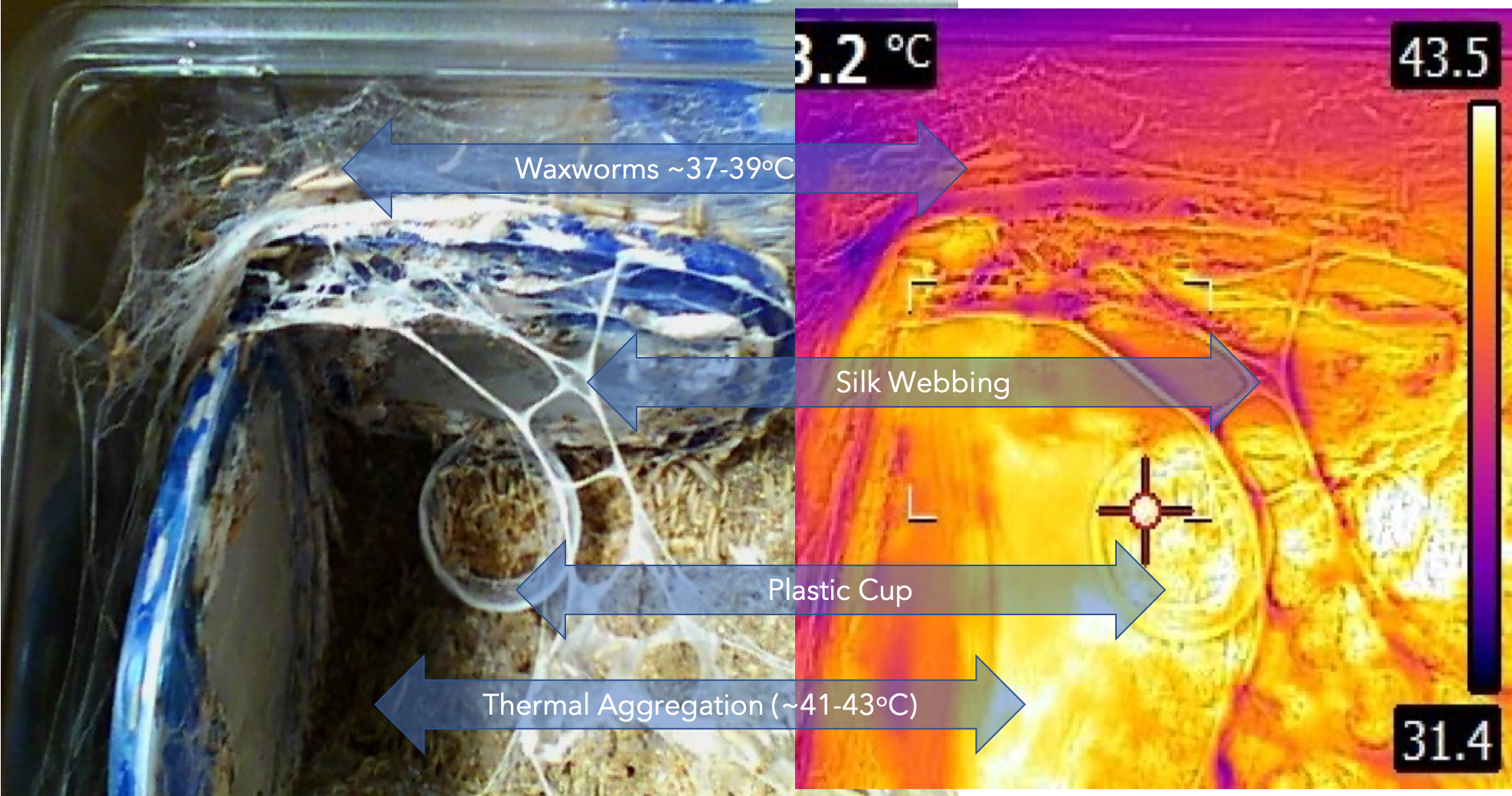
These interactions are illustrated in the images taken from my waxworm colony where I used a thermal imaging camera to capture the heat (thermal) gradient produced by the waxworms in this container.
Left: visual light image of waxworms and their architecture: right: infra-red image of the same colony space, showing the thermal gradients associated with aggregations of waxworms and the silken structures that the waxworms generate. This is the subject of a paper that I will soon be publishing in the journal INSECTS.
In the study of the colony described here, I have also included (besides the thermal profile) the CO2 and O2 gradient that results from the waxworms’ behavior and metabolism. The example, taken from waxworm cultivation in my research program, is typical of the kinds of multi-dimensional dynamics that apply to ALL insect rearing systems. My whole point in this and related discussions is to get students of insect rearing to recognise the complexities of their rearing systems components. It is further intended to awaken students’ appreciation for knowing the nature of (the science of) these many factors and their interactions.
This approach is not simple or easy; but it is very powerful in establishing and maintaining healthy, productive colonies!
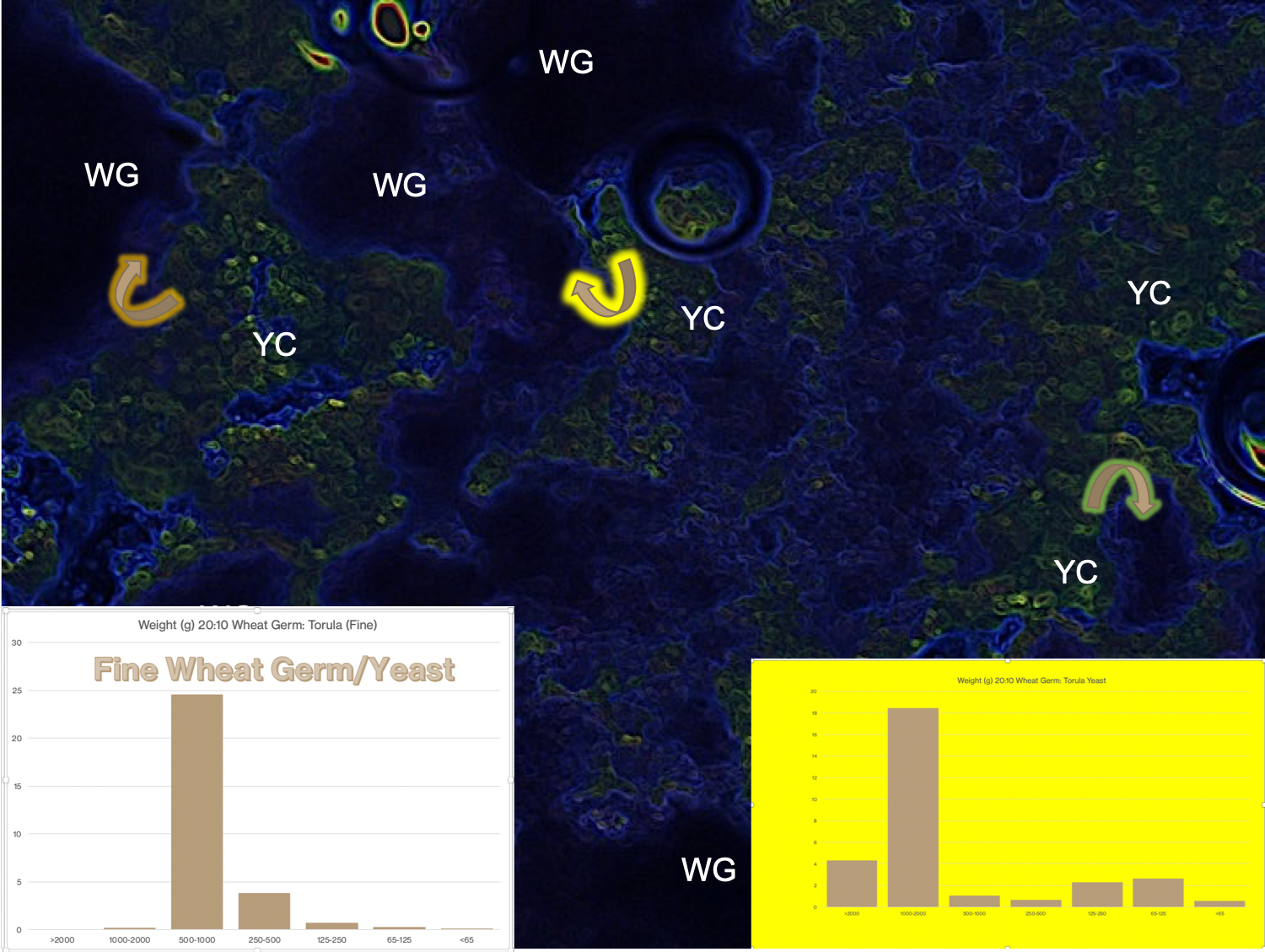
In the previous post, I mentioned the concept of granulometry as part of the approach to understanding matrix relationships–in this case, the particle-particle interactions.
An approach to granulometry hybridising visual/image-based analysis and particle size analysis by sieve separation techniques.
The above figure is a representation of the image in the previous discussion where the following modifications were made to the original figure: 1) the color pattern was changed in an image processing step performed in Microsoft PowerPoint. The technique changes colors and adds emphasis and contrast to the different portions of the image; 2) portions of the figure that contain wheat germ (WG) and yeast concentrations (YC) were marked, and some of the suggested interactions were depicted with the arrows; 3) the insets represent two different kinds of granulometric analyses where the coarse diet solids (wheat germ and torula yeast) on the lower right and fine particles on the lower left were separated with a set of mechanised sifting screens. Particles within each size range were weighed and the size distributions graphed with the bar graphs. The bar graphs represent these particle sizes: > 2000 microns; 1000-2000 microns; 500-1000 microns; 250-500 microns; 125-250 microns; 65-125 microns; and < 65 microns. The different sizes of particles between the coarse and fine particles were produced by using mixtures of wheat germ (20 g) and torula yeast (10 g) of non-milled solids vs. the same proportions of the solids that had been run through a cutting mill.
In association with the differences in particle size, the diet made from the coarse particles had a firmness (gel strength) that was about 1.5 times as great as that of the gel made with fine particles. This is an example of the kinds of effects that matrix interactions can influence in various artificial diets. Part of our current research in the Insect Rearing Education and Research Program at NCSU is to discover the various kinds of matrix characteristics that arise from use of various diet materials and diet-processes. A central line of inquiry deals with the physicochemical characteristics of different factors in diet production.
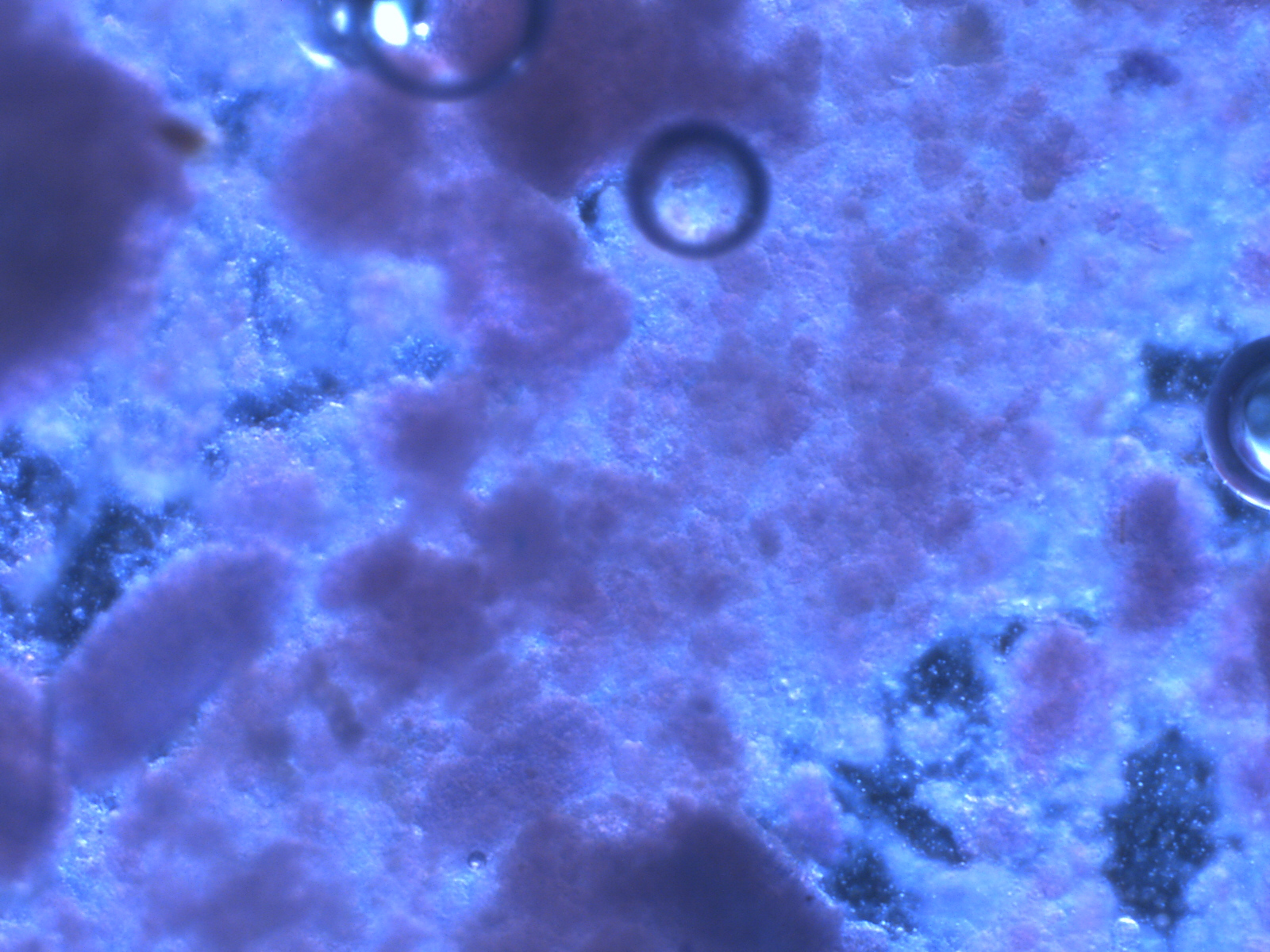
In yesterday’s post, I discussed the matrix concept in insect diets. I used an example of a visual treatment of a diet matrix using an image of a stained wheat germ-based diet viewed and photographed under about 20 x magnification in a video microscopy system. The basic image looked like this:
Section of a wheat germ and torula yeast diet stained with Eosin and Methylene Blue, then visualised with a stereomicroscope at 20 X.Macro-view of hornworm neonates (Manduca sexta) feeding on the wheat germ diet.
In the above images, the lower image shows a 1 x 3 cm section of diet with hornworms consuming the diet. The neonates (newly-hatched 1st instar larvae) depend upon finding all the nutrients they require within a small area of the diet where all lipids, protein, carbohydrates, minerals, and vitamins must be present in adequate amounts to support healthy growth. The upper image shows the granular nature of the diet whose particles (which are suspended in an agar or carrageenan framework) have an organization that can be thought of as conforming to a network, matrix relationship. This relationship (the granulometery) contributes to the potential for structural integrity, resulting in diet consistency and texture. There is also a great deal of potential chemical interaction such as lipid-binding by lipoproteins, carbohydrate-protein interactions, enzymatic and oxidative reactions, diffusion of solutes or nano-particles that are involved in the diet’s taste/texture qualities (palatability), the availability of nutrients within the reach of the insect’s mouthparts; then upon ingestion, the particles and solutes must be available for digestion and absorption. Finally, the arrangement of the particles and other diet components must lend itself to the diet’r retention of palatability, nutritional value, and bioavailability in a framework that we call stability. All the physicochemical characteristics and interactions constitute the diet’s MATRIX.
In these images, there is an implicit matrix organization that dictates or commands the nature of the diets in question. To better understand this matrix/organization feature, we can examine the following diagram:
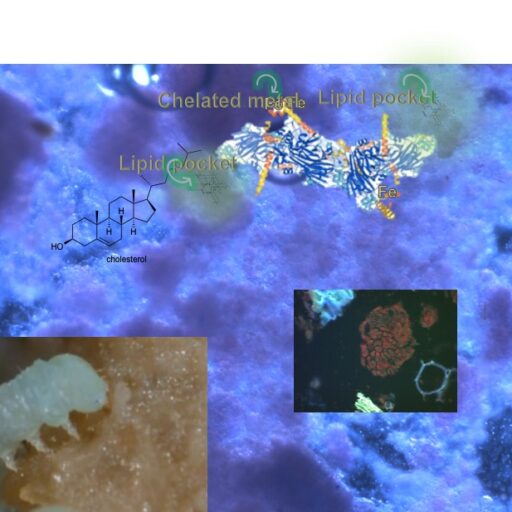
Diagram of scale in an insect diet. This image includes a background that illustrates a micrograph of the wheat germ diet (a 2 mm x 2 mm portion with the blue/pink stained diet to help illustrate the particle arrangement = granulometry) with inset images that show proportions. The lower left image shows a 70 mm x 70 mm section of a rearing container with Manduca sexta larvae feeding on the wheat germ diet. The lower right inset image shows a 7 x 7 mm portion of diet with an M. sexta neonate feeding.
In the above diagram (partly explained in the image caption), the various levels of organization are shown from macro- to nano levels. The inset on the right-central part of the diagram is a fluorescent micrograph of the diet that was stained with the lipid stain, Nile Red, which shows the lipids in a 20 micron particle of wheat germ. Other fragments in this insect (against the black background) are brightly-coloured carbohydrates, which stain blueish with Nile Red. The upper central portion is a diagram of a large lipoprotein molecule of about 25 nm x 5 nm, and on the left central portion a sterol molecule is depicted as a less than 1 nm structure. The lipoprotein molecule contains hydrophobic pockets (the greenish structures on the left and right ends of the molecular structure). It is the hydrophobic pockets in lipoproteins that bind with and carry lipids (= lipophorins). Throughout this diagram, it is evident that there are micro-relationships on the physicochemical level of organization, and these relationships lead to degrees of stability, nutrient bioavailability, and taste (palatability). An important part of this relationship is in the interactions between particles and liquid interfaces, and we will further explore these in relationships that we study with granulometry.